Université de Liège - Centre de l'Oxygène, Recherche et Développement (CORD)
Université de Liège - Centre de l'Oxygène, Recherche et Développement (CORD) Université de Liège - Centre de l'Oxygène, Recherche et Développement (CORD)
L'Oxygène et la Vie: Tome II - L'Oxygène en Pathologie des Mammifères
Mitochondries et métabolisme de l'oxygène
Carol Deby
La privation d'oxygène est supportée de manières diverses par les êtres vivants aérobies. Certaines cellules, comme les chondrocytes constituant le tissu cartilagineux, vivent dans une concentration d'oxygène correspondant à 5%.
Mais l'oxygène est indispensable à la vie et sa privation entraîne la mort, avec une rapidité dépendant de la nature des cellules.
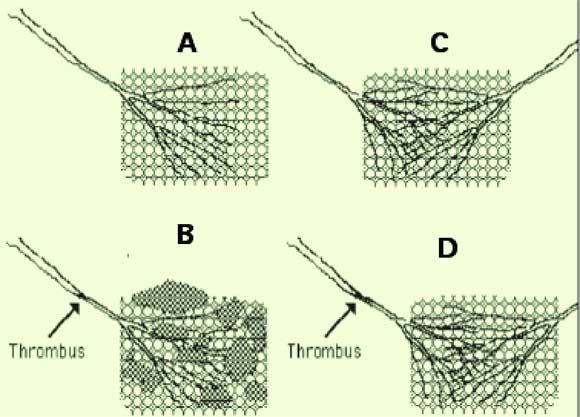
| Fig. 1 : Dans la partie gauche de la figure, les tissus (schématisés par un grillage) ne sont irrigués que par une seule artère (A); l’obstruction de celle-ci par un thrombus entraîne la nécrose du tissu qu’elle alimentait (B). Dans la partie droite, deux artères se partagent l’irrigation du tissu (C). La thrombose d’une des artères est compensée par un débit de sang accru dans l’autre artère (D). Le premier cas est un exemple de circulation terminale, le deuxième de circulation anastomosée. |
Mécanisme de la mort par anoxie.
Durant les premières décennies de la biologie (avant 1900), la mort par privation d’oxygène semblait un fait tellement avéré qu’on ne pensait pas qu’il y eut des mécanismes mettant en cause des phénomènes chimiques et, encore moins, des réactions de biologie moléculaire. Ce n’est que vers les années trente avec la découverte des cytochromes, du cycle de Krebs et du rôle d’accumulateur d’énergie que constitue l’ATP, que l’on se rendit compte que, pour exister, les cellules produisent de l’énergie chimique d’une manière analogue à la combustion, ce qui demande de l’oxygène. Mais cette combustion ne peut être instantanée, comme lors de l’allumage d’un feu; elle doit s’effectuer par petites étapes: « La cellule a besoin de petite monnaie » avait déclaré le grand Michaélis.
La fin des années cinquante vit le développement des techniques permettant l’observation des tissus et cellules en microscopie électronique, en même temps que se développait l’ultracentrifugation fractionnée. On put voir, agrandies des dizaines de milliers de fois, ces organelles (ou organites) devinées en microscopie optique. On put les isoler, les faire travailler et on reconnut que toute la machinerie productrice d’énergie est localisée dans ces espèces de saucisses d’où sortent les molécules d’adénosine triphosphate (ATP); ces accumulateurs énergétiques cèdent leurs calories dans le cytosol là où se déroulent les réactions endothermiques (ou endergoniques); l’ATP est le combustible des moteurs qui permettent la contraction musculaire, le déplacement d’une cellule mobile, le fonctionnement des pompes à ions, etc... L’ATP est produit par les mitochondries qui occupent ainsi une position clé en physiologie.
La libération d’énergie résultant de la réduction de molécules d’oxygène en eau est une réaction hautement exothermique qui est permise par l’arrivée continue d’électrons arrachés à des substrats organiques (cycle de Krebs). Ces électrons sont apportés par une chaîne de transport le long de laquelle ils perdent progressivement leur énergie électrochimique, utilisée pour transformer l’acide adénosine diphosphorique (ADP) en acide adénosine triphosphorique (ATP). Lorsque les électrons appauvris en énergie arrivent en bout de chaîne et rencontrent les molécules d’oxygène, le dégagement de calories est fortement affaibli: il ne se produit qu’un faible dégagement de chaleur. Le lecteur peu familiarisé avec ces notions électrochimiques peut comparer ces pertes progressives d’énergie à la chute d’un homme tombant de cinq étages et transformant, en peu de secondes, en énergie cinétique l’énergie potentielle qu’il avait acquise en montant au cinquième étage. Les résultats mécaniques qui produisent un polytraumatisme dans le cas de la défenestration seront tout à fait différents si le sujet descend les cinq étages en empruntant l’escalier. La transformation de l’énergie potentielle sera beaucoup plus longue et le sujet arrivera sain et sauf au rez-de–chaussée.
Ces organelles produisent continuellement des radicaux libres et des espèces oxygénées activées (ROS). Lors de la reperfusion, cette production augmente et, avec des troubles du transport du calcium, menace l’intégrité mitochondriale et provoque notamment l’ouverture d’un pore de la membrane externe de l’organelle. Ce pore, ouvert, laisse sortir dans le cytosol certaines protéines déclenchant une cascade de réactions aboutissant à l’activation, dans le noyau cellulaire, de DNAases qui vont morceler l'acide désoxyribonucléique (ADN ou DNA:desoxyribonucleic acid): c’est la mort cellulaire par apoptose.
Mitochondries et métabolisme de l’oxygène
Ces organelles sont :
• Première partie : Mitochondries et oxygénation
Sommaire
Chapitre 1 : Généralités
Chapitre 2 : Phosphorylation oxydative
Chapitre 3 : Régulation de la chaîne de transport des électrons et de la phosphorylation oxydative
Chapitre 4 : Phénomènes redox mitochondriaux
Chapitre 5 : Ions et canaux membranaires mitochondriaux
Chapitre 6 : Production de ROS par les mitochondries
Chapitre 7 : Rôle des mitochondries dans l’apoptose
Chapitre 8 : Protection anti-ROS mitochondriale
Bibliographie
Glossaire
• Deuxième partie : Troubles de l’oxygénation et mitochondries
Sommaire
Chapitre 1 : No-reflow phenomenon
Chapitre 2 : Anoxie-réoxygénation
Chapitre 3 : Ischémie–reperfusion
Chapitre 4 : Ischémie-reperfusion: traitements
Chapitre 5 : Synopsis du rôle des ROS dans les affections cardiaques
Bibliographie
Glossaire
 |
 |
 |